

【Nat. Neurosci.】傲雪凌霜!马普所揭示机体应对负面情绪的环路机制
正如傲雪凌霜的松柏,生活中面对困境时,我们披荆斩棘,乘风破浪,战胜艰难险阻,一步步实现人生目标。在此过程中,我们的机体感知负面状态并作出适应性调整。除人类以外,这种适应性功能对其它生物的生死存亡同样至关重要。
 图片来源:Liz Burton个人主页
图片来源:Liz Burton个人主页
过去研究表明,岛叶皮质后部(posterior insular cortex, pIC)整合机体内、外部环境信息,处理多种功能[1-5]。但是pIC如何表征身体内部状态以及如何介导相关适应性行为,我们尚未可知。
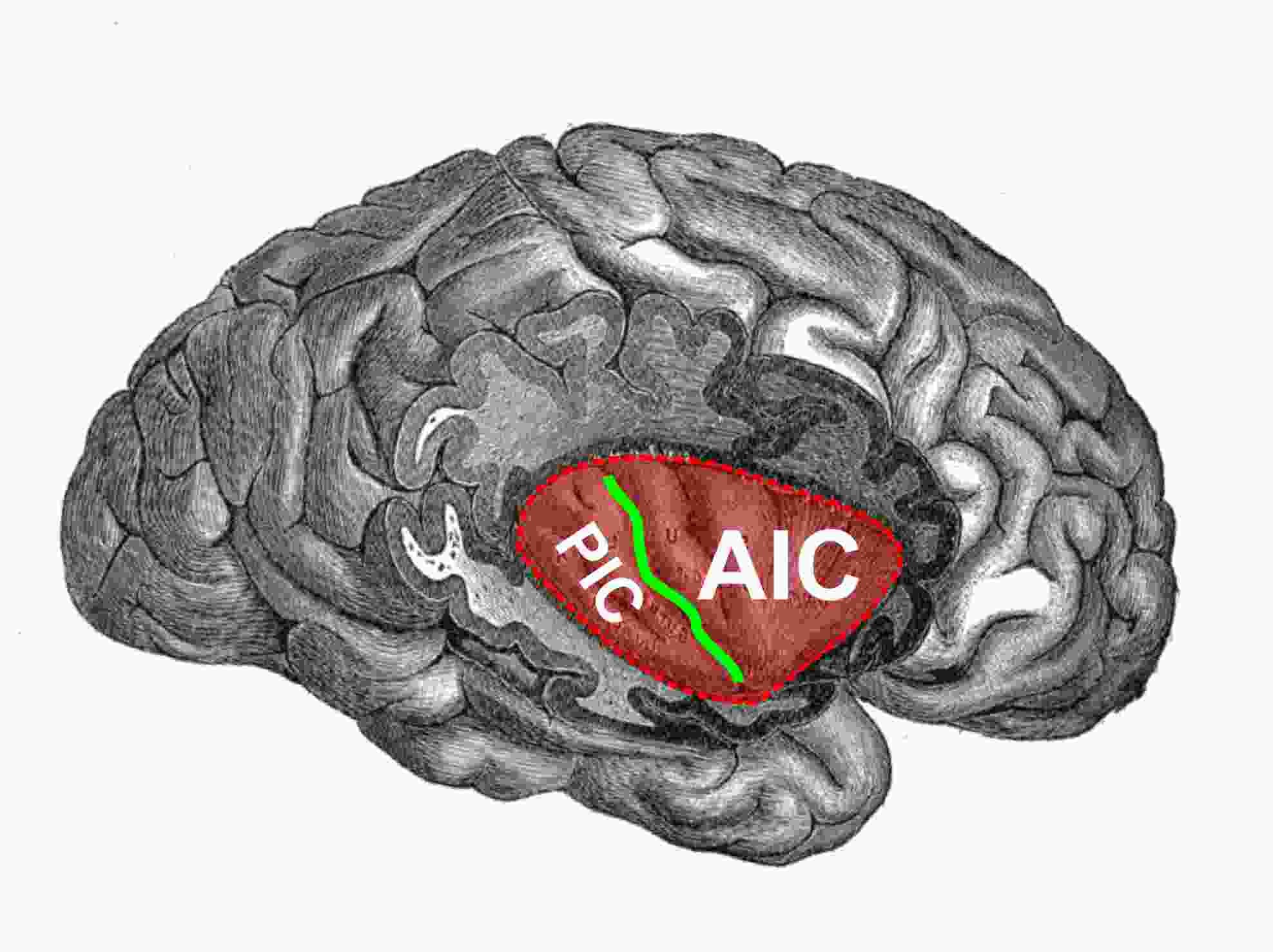
2019年8月27日,《Nature Neuroscience》杂志在线刊登了德国马克思普朗克神经生物学研究所Nadine Gogolla研究组的最新重要工作[6],他们发现pIC通过不同下游编码机体对抗负面状态引起的多种行为。这项研究揭示了介导负面状态相关适应性行为的神经环路机制,极大提高了人们在负面状态相关神经疾病领域的认知。
 Nadine Gogolla, PhD 图片来源:Max Planck Institute of Neurobiology
Nadine Gogolla, PhD 图片来源:Max Planck Institute of Neurobiology
为探究pIC编码的功能,作者在小鼠pIC双侧注射AAV-CaMKIIα-ChR2或AAV-CaMKIIα-eNpHR3.0并植入光纤(图1a),发现光激活pIC引起梳毛、下颌运动、颤栗和躲避行为(图1b-d)。在麻醉小鼠中,光激活pIC增加呼吸频率,光抑制pIC无影响(图1e)。以上结果表明激活pIC产生一种负面信号,引起小鼠产生防御性反应。
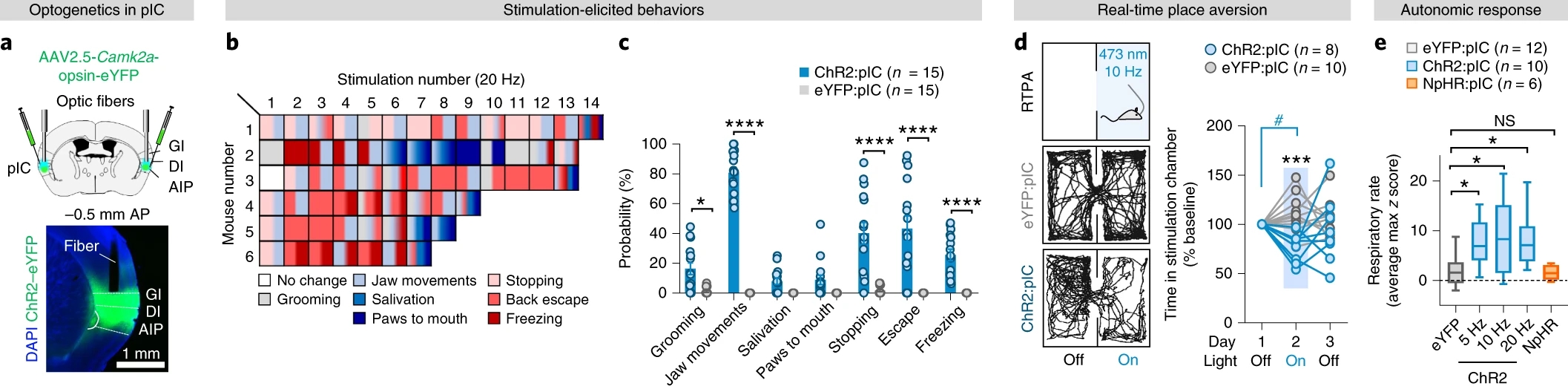 图1光激活pIC诱发防御行为和自主神经反应
图1光激活pIC诱发防御行为和自主神经反应
为探究何种状态与刺激引起pIC激活,作者使用光纤钙信号记录方法,在小鼠pIC中注射AAV-CaMKIIα-GCaMP6s(图2a),将小鼠置于高架十字迷宫(EPM)或高架O迷宫(EZM)中。在两种高架迷宫中,小鼠在开放臂的焦虑水平更高。他们发现,小鼠在开放臂中pIC活性较低,且在开放臂中逗留时间更长(焦虑水平更高)的小鼠在开放臂中pIC活性更低(图2b-d)。此外,小鼠撤离开放臂时,pIC活性升高;小鼠接近开放臂时,pIC活性降低(图2e-h)。
为研究pIC是否调控焦虑行为,作者在小鼠pIC中双侧注射AAV-CaMKIIα-ChR2或AAV-CaMKIIα-eNpHR3.0并植入光纤,发现光抑制pIC增加开放臂探索,光激活pIC减少开放臂探索(图2i-l)。
以上结果表明pIC活性与小鼠焦虑水平呈正相关,操控pIC神经元可充分改变小鼠焦虑水平。
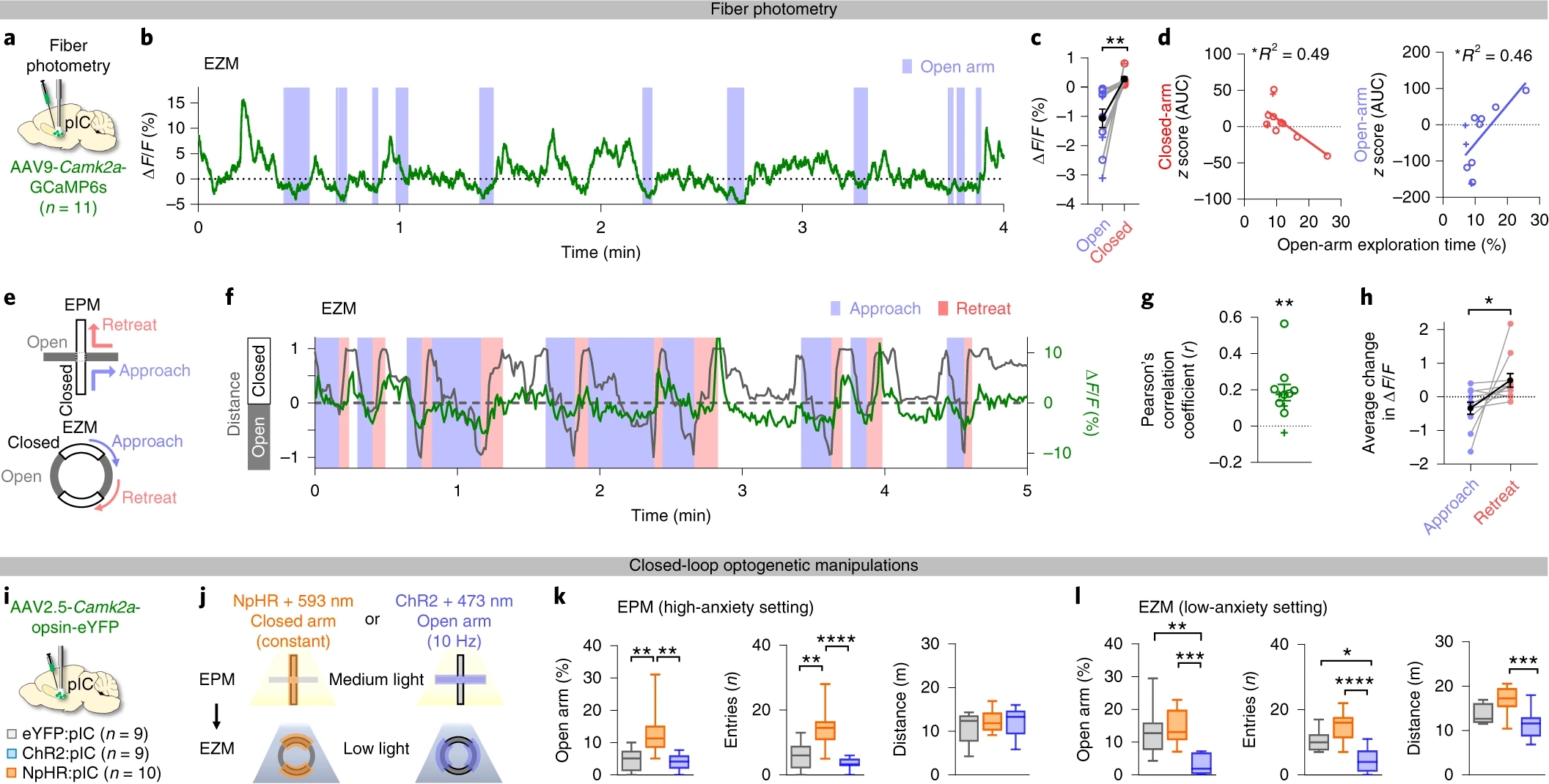 图2 pIC参与并调控焦虑行为
图2 pIC参与并调控焦虑行为
为进一步探索pIC是否参与焦虑状态的产生,作者研究短暂操控pIC神经元活性是否产持续性行为学效应。他们继续使用上文中光遗传学方法,给予小鼠20Hz光刺激10次,每次持续1秒,然后将小鼠置于EPM中,发现小鼠在开放臂中逗留时间显著减少;NpHR3.0实验中,作者先使用足底电击范式提升小鼠焦虑水平,在此过程中光抑制pIC,再将小鼠置于EPM中,发现小鼠在开放臂中逗留时间显著增加(图3a-d),表明短暂操控pIC神经元活性可以引起小鼠焦虑水平的持续性变化。
为排除NpHR3.0实验中恐惧情绪带来的影响,作者在场景恐惧范式足底电击或场景回忆过程中光抑制pIC,发现小鼠恐惧行为及恐惧记忆并不受影响(图3e-f)。
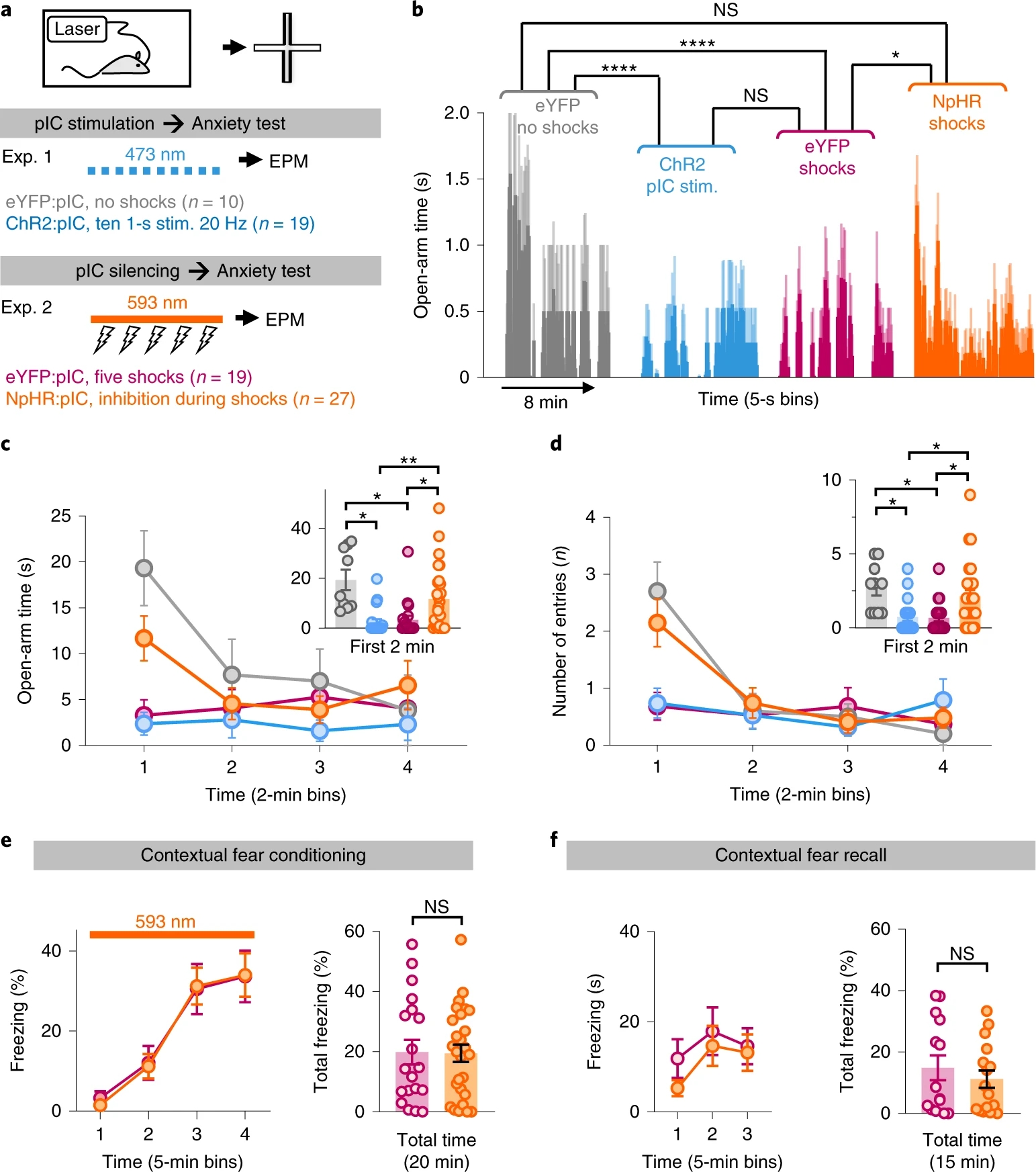 图3 短暂操控pIC引起焦虑水平的持续性变化
图3 短暂操控pIC引起焦虑水平的持续性变化
上文已知,pIC介导焦虑等负面情绪。但是,我们并不知清楚负面情绪如何在pIC中产生与处理。过去研究表明,岛叶皮层接受多信息流输入[2],于是作者探究不同情绪性刺激如何由pIC处理。
他们引入双光子钙成像方法,在小鼠pIC中注射AAV-CaMKIIα-GCaMP6s,给予小鼠尾部电击、糖水或奎宁溶液以表征痛觉、奖赏性、厌恶性刺激,发现28% pIC神经元被尾部电击激活,49%被奎宁溶液激活,10%被糖水激活,7%被恐惧性颤栗激活(图4a-e)。
接着,为探究pIC在单细胞水平如何表征焦虑状态,作者比较小鼠在条件恐惧学习后(高焦虑状态)与条件恐惧消退后(低焦虑状态)pIC神经元的活性,发现前者显著高于后者(图4g)。
再然后,作者探究pIC在单细胞水平如何表征机体厌恶性状态,他们通过腹腔注射氯化锂溶液或水剥夺处理以引起小鼠身体不适,发现大量pIC神经元被这两种负面状态激活(图4g)。
统计结果显示,23.5%神经元被多种刺激激活;超过50%神经元被焦虑状态激活,12.2%神经元被焦虑状态与痛觉刺激激活;超过99%神经元参与身体状态检测,其中75%参与多种厌恶性状态检测(图4f, h)。
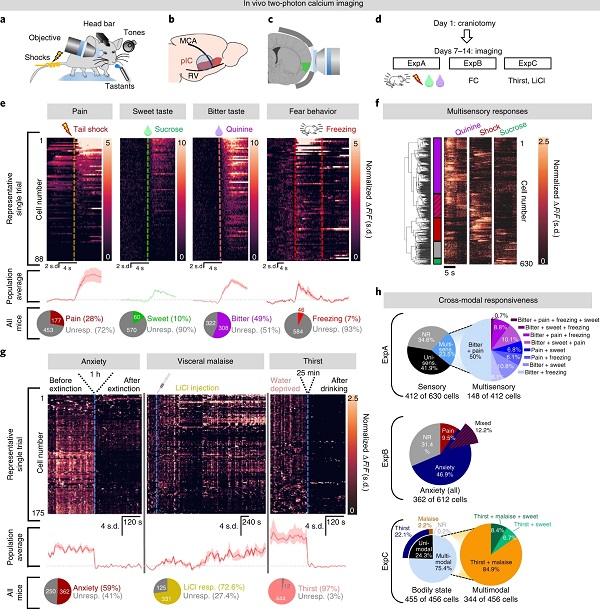 图4 pIC中厌恶性刺激与状态的单细胞表征
图4 pIC中厌恶性刺激与状态的单细胞表征
 图5 pIC的上、下游脑区
图5 pIC的上、下游脑区
过去的研究表明,中央杏仁核(CeA)与伏隔核核部(NAcC)参与并调控焦虑、厌恶与奖赏等功能[7-9],且这两个脑区为pIC的主要下游(图5c),于是作者推测此二脑区介导pIC参与的功能。
为研究投射到CeA与NAcC的pIC神经元是否为同一集群,作者在小鼠CeA与NAcC中分别注射红、绿色CTB(图6a-b),发现pIC中CeA与NAcC的上游神经元极少共标,其中CeA上游神经元分布靠后,NAcC上游神经元分布靠前(图6c-f)。他们又在CaMKIIα-Cre小鼠bregma -0.5 mm位置注射AAV-DIO-eYFP,找到了CeA与NAcC中eYFP阳性神经纤维最密集的区域(图6g-j),为下文中的光纤埋植位点提供最优坐标。
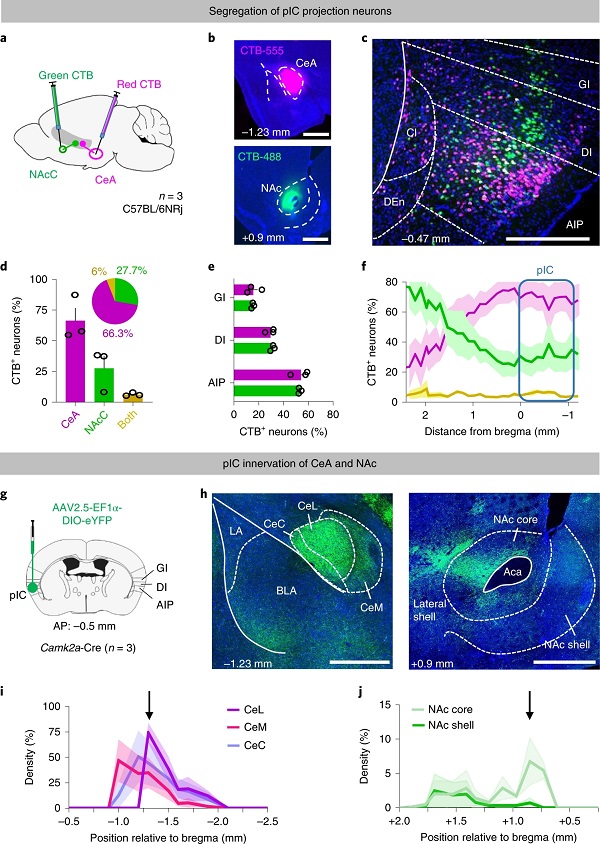 图6 投射到CeA 与NAcC 的pIC神经元分属不同亚群
图6 投射到CeA 与NAcC 的pIC神经元分属不同亚群
为探究pIC→CeA和pIC→NAcC环路的功能,作者在小鼠pIC中注射AAV-CaMKIIα-ChR2或AAV-CaMKIIα-eNpHR3.0,在CeA或NAcC植入光纤(图7a-b),发现光激活pIC→CeA环路增加厌恶性行为、躲避行为、呼吸频率以及焦虑行为,而光激活pIC→NAcC环路无此现象。此外,光抑制pIC→CeA环路减少焦虑行为(图7c-n)。
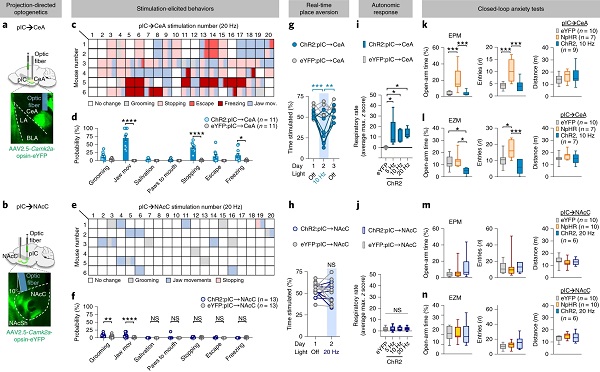 图7 pIC→CeA介导厌恶、焦虑行为与自主神经反应
图7 pIC→CeA介导厌恶、焦虑行为与自主神经反应
过去研究表明,CeA与NAcC均参与满足行为[7, 9, 10]。满足行为是一种高动机行为,例如在危险的条件下获取奖赏,“贪夫徇财,烈士徇名”是其典型案例。接下来,作者探究CeA与NAcC是否介导pIC调控满足行为。
他们使用与上文相同的光遗传学操控方法,发现光激活两条环路均会显著减少小鼠饮用糖水的时程,而光激活、抑制两条环路不影响小鼠对糖水的偏好性(图8a-e)。与之相似,光激活两条环路均会中断小鼠进食,减少其进食量(图8f-h)。此外,光激活pIC→CeA而非pIC→NAcC环路会中断小鼠社交过程,但小鼠社交总时间维持不变(图8i-k),表明pIC→CeA环路并不影响小鼠社交动机。
厌恶性状态引起饥饿小鼠厌食,最后,作者探究两条环路在此过程中的作用。他们发现,光抑制pIC→NAcC环路后,身体不适或焦虑气味引起的厌食现象有所缓解,而光抑制pIC→CeA环路无任何影响(图8l-r)。
综上所述,pIC→CeA环路介导厌恶、焦虑行为与自主神经反应,pIC→NAcC介导满足行为。
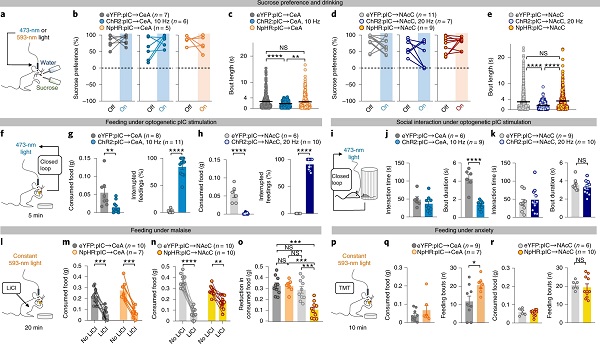 图8 pIC→NAcC介导满足行为
图8 pIC→NAcC介导满足行为
负面状态的感知与其适应性行为对各类生物的生存十分重要,但是究其相关神经环路机制,我们知之甚少。本篇文章结合光纤钙信号记录、光遗传学、双光子钙成像、与神经元顺、逆行示踪等方法,发现pIC编码负面状态引起的各类行为,其中pIC→CeA环路介导厌恶、焦虑行为及其相关自主神经反应,pIC→NAcC环路介导满足行为。这项研究阐释了机体应对负面情绪的神经环路机制,为临床治疗焦虑症等负面情绪相关疾病提供有力支持!
和元上海一直关注神经科学领域的重大研究进展,为神经生理、病理研究提供最新工具和研究方案,助力临床转化和基因治疗!
参考文献

扫一扫,反馈当前页面
和元生物
