在中枢神经系统中,突触的传递效能会随着神经活动的变化而改变,这称之为“突触可塑性”,被认为是大脑高级功能学习与记忆的细胞基础。在大脑海马CA1兴奋性神经元中,位于树突棘(Dendritic spines, 是神经元树突上的微小突起,是兴奋性突触的主要突触后位点)突触后膜的AMPA型谷氨酸受体数量的增加和功能的增强介导了突触长时程增强(long-term synaptic potentiation,LTP)效应,这被称为是突触可塑性的经典形式。
神经元间突触的信号传递与树突棘的可塑性密切相关。神经信号如何诱导树突棘产生功能可塑性是一个重要而有趣的神经细胞生物学问题。以往大量的研究聚焦于蛋白质分子在突触可塑性中的功能与作用机制,而对于细胞中另一类重要的生物大分子―脂质分子鲜有涉猎。
中国科学院遗传与发育生物学研究所刘佳佳研究员团队借助脂质组学、活细胞成像、细胞生物学技术、RNAi(RNA干扰)技术、全细胞膜片钳记录、行为学范式等多个技术手段发现位于兴奋性神经元树突胞膜上PI4P的代谢调控在活动依赖的AMPAR膜转运过程中以及在LTP和学习记忆中发挥重要作用,并揭示了活动诱导的细胞内钙离子浓度上升促进了PI4KIIIa向细胞膜表面募集,进而促进PI4P的合成过程。相关研究成果于2022年3月1日刊发在《Cell Reports》上,标题为“Activity-dependent PI4P synthesis by PI4KIIIα regulates long-term synaptic potentiation”。

首先,为了明确磷脂酰肌醇4-磷酸(Phosphatidylinositol 4-phosphate, PI4P)在神经元中的亚细胞分布,作者借助免疫荧光技术发现,海马神经元细胞膜中PI4P信号强度明显高于Neuro-2a(小鼠脑神经瘤细胞)、NIH/3T3(小鼠成纤维细胞)和 HeLa(上皮细胞)。定量分析发现,PI4P在轴突胞膜中比树突更丰富(图1)。
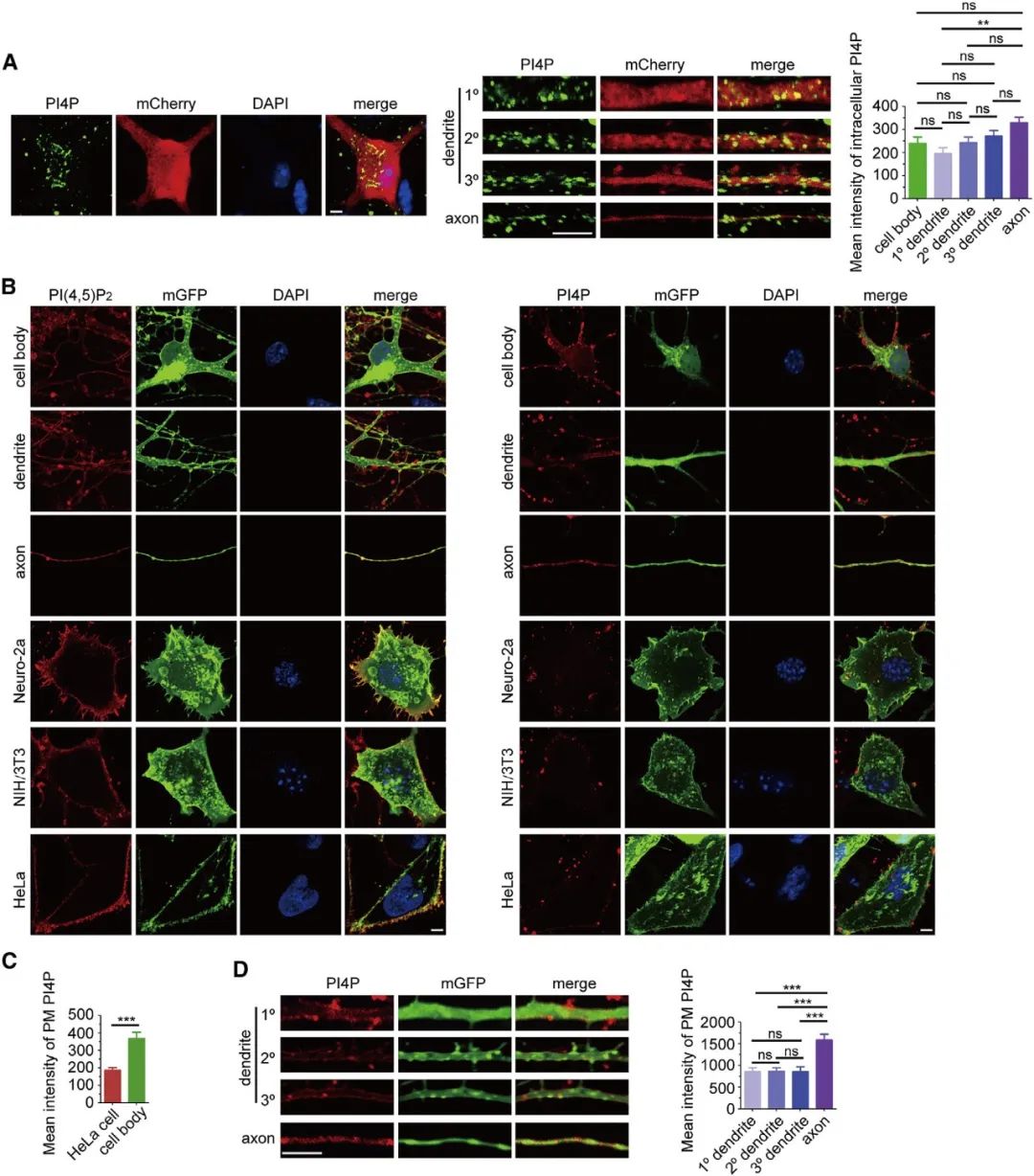
图1 PI4P在小鼠海马神经元中的亚细胞分布
2.树突胞膜中的PI4P水平在LTP过程中发生动态可逆变化
在中枢神经系统中,位于树突棘的突触后位点接收了大部分兴奋性输入。为了探索PI4P在突触传递和突触可塑性中的作用,作者首先研究了PI4P在树突中的亚细胞分布和含量是否会随着神经活动发生动态变化。在体外培养的海马神经元中,研究者借助NMDA受体的共激动剂(co-agonist)甘氨酸化学诱导细胞的长时程增强(long-term synaptic potentiation,LTP)效应。结果发现,树突的PI4P信号强度受神经元活动的调节,其水平在LTP期间经历动态可逆变化(图2)。
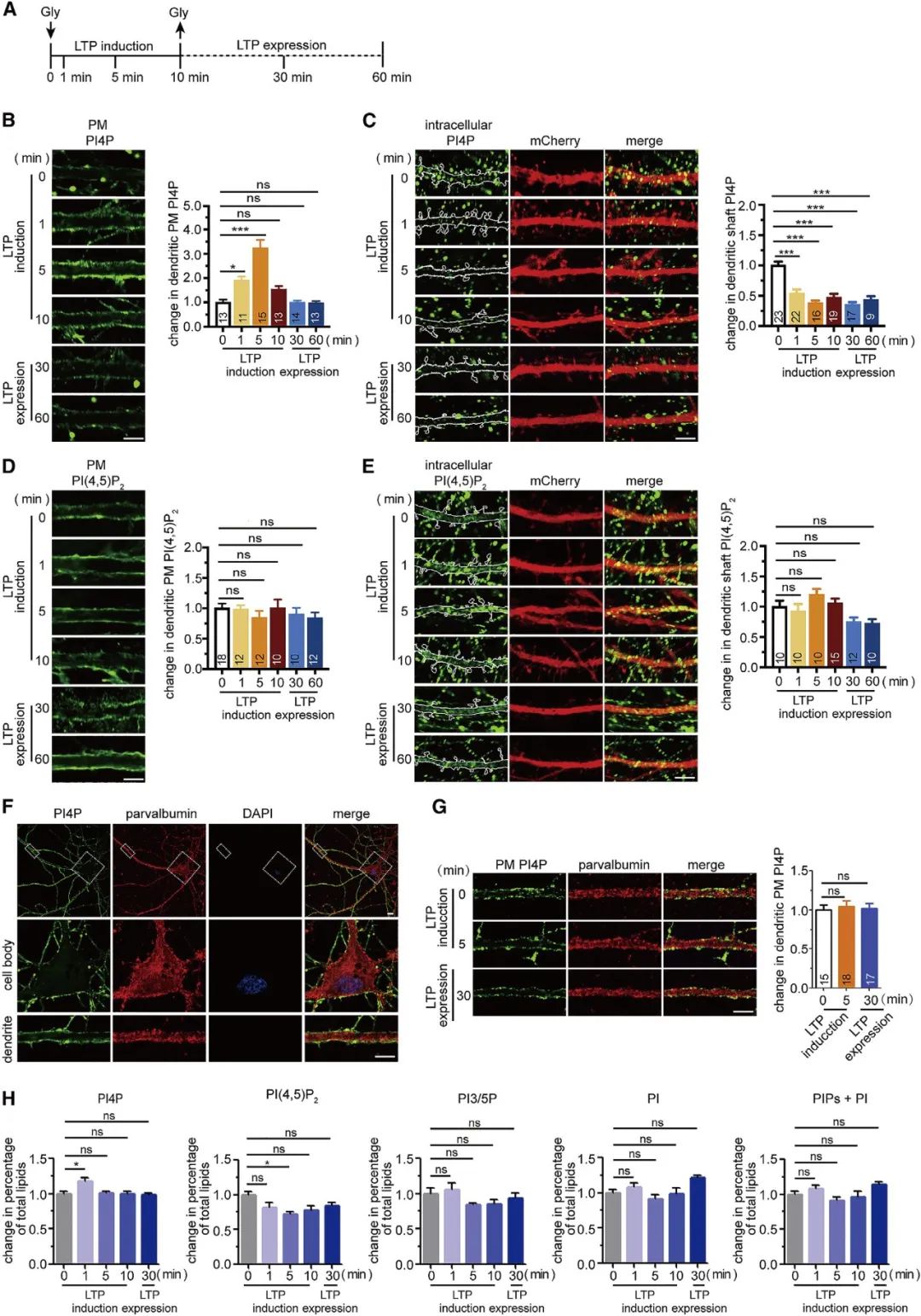
图2 树突胞膜中的PI4P水平在LTP过程中发生动态可逆变化
基于上述结果,作者假设PI4P参与了活动依赖的突触后成分的膜转运(如AMPA将受体转运至突触位点上),为了验证该假说,首先需要调控PI4P在胞膜或细胞内膜上的含量。以PI为底物,共有四种激酶可以将其磷酸化为产物PI4P,其中PI4KIIIa被认为参与了PI4P在胞膜中的合成和维持。借助化学抑制剂或AAV介导的shRNA表达载体(pAKD-CMV-bGlobin-eGFP-H1-PI4KIIIa shRNA),研究者在海马神经元中抑制PI4KIIIa的活性或表达。结果显示,抑制PI4KIIIa可导致树突胞膜中PI4P缺失。进一步地,在化学诱导LTP的过程中急性给予PI4KIIIa抑制剂可以阻止胞膜上PI4P水平的瞬时性上升。这些结果提示了,在神经元树突胞膜上,PI4KIIIa既与PI4P静息状态下的含量相关,也参与了活动依赖的PI4P合成过程。(图3)。
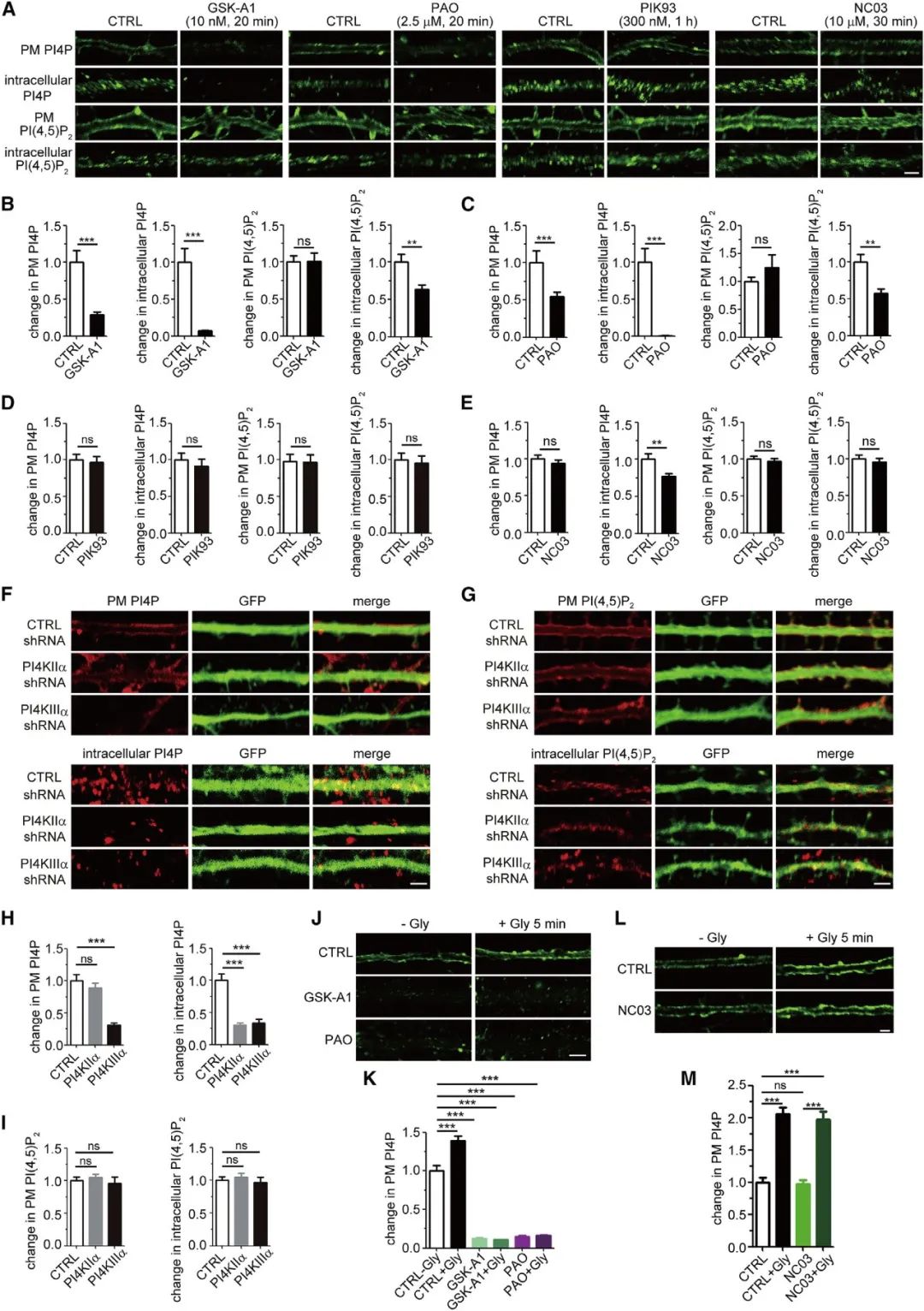
图3 PI4KIIIa参与了活动依赖的PI4P合成过程
接下来,研究人员验证了PI4P在活动依赖的AMPAR膜转运过程中的重要作用,同时,借助免疫染色发现,树突上大多数AMPAR转运体并不富含 PI4P,提示了细胞膜上的PI4P是活动依赖的AMPAR囊泡胞外转运所必需的。
随后,研究人员进一步发现CICR(钙诱导的钙释放)促进了PI4KIIIa在LTP期间从细胞质到细胞膜表面募集,进而介导了PI4P的合成,促进AMPAR亚基GluA1在细胞膜上的运输(图4)。
上述结果共同提示了产生LTP的海马神经元中,细胞内钙离子浓度上升促进了PI4KIIIa向细胞膜表面募集,进而促进PI4P的合成过程。
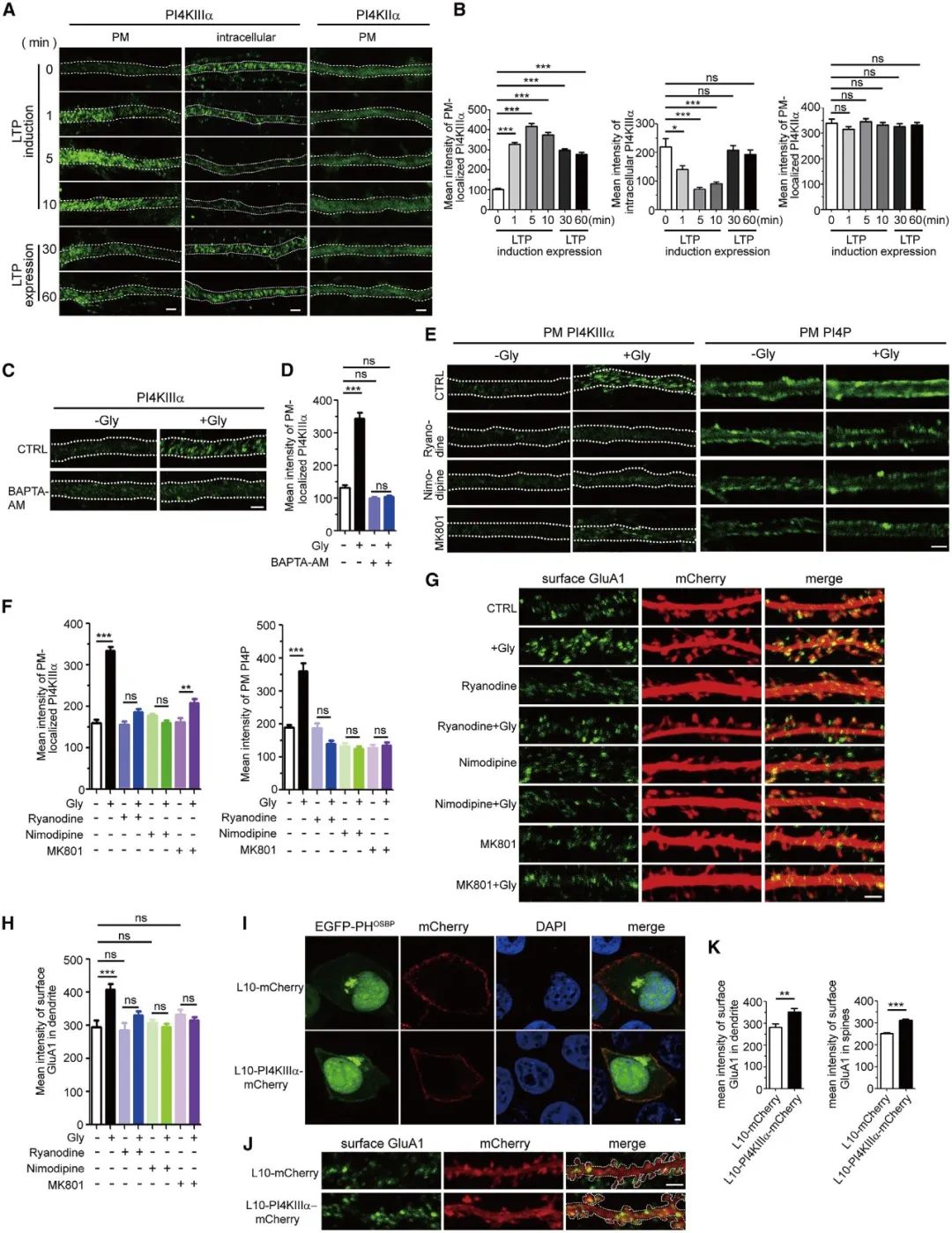
图4 PI4KIIIa定位树突质膜表面,并在LTP诱导下催化合成PI4P
4.沉默PI4KIIIa表达可抑制LTP和长期记忆
为了研究干扰PI4KIIa或PI4KIIIa是否会影响神经元的突触传递,研究人员在成年小鼠海马CA1区注射pAKD-CMV-bGlobin-eGFP-H1-PI4KIIa shRNA或pAKD-CMV-bGlobin-eGFP-H1-PI4KIIIa shRNA病毒。全细胞膜片钳记录的结果显示,在海马神经元中沉默PI4KIIa或PI4KIIIa沉默导致了AMPAR介导的兴奋性突触后电流(EPSC)发生了轻度下降(图5A-F)。
为了研究沉默PI4KIIa或PI4KIIIa是否会影响突触可塑性,研究人员首先在E14.5的小鼠胚胎中侧脑室注射PI4KIIa或PI4KIIIa的shRNA质粒,然后在P14-21之间制备了小鼠的急性脑切片并进行电生理记录。LTP分析显示,沉默PI4KIIIa不仅显著降低了LTP的幅度(magnitude),并且破坏了CA1神经元中LTP的维持(图5M,N)。这些数据提示了PI4KIIIa依赖的PI4P合成对于LTP期间的树突棘的结构和功能可塑性至关重要。
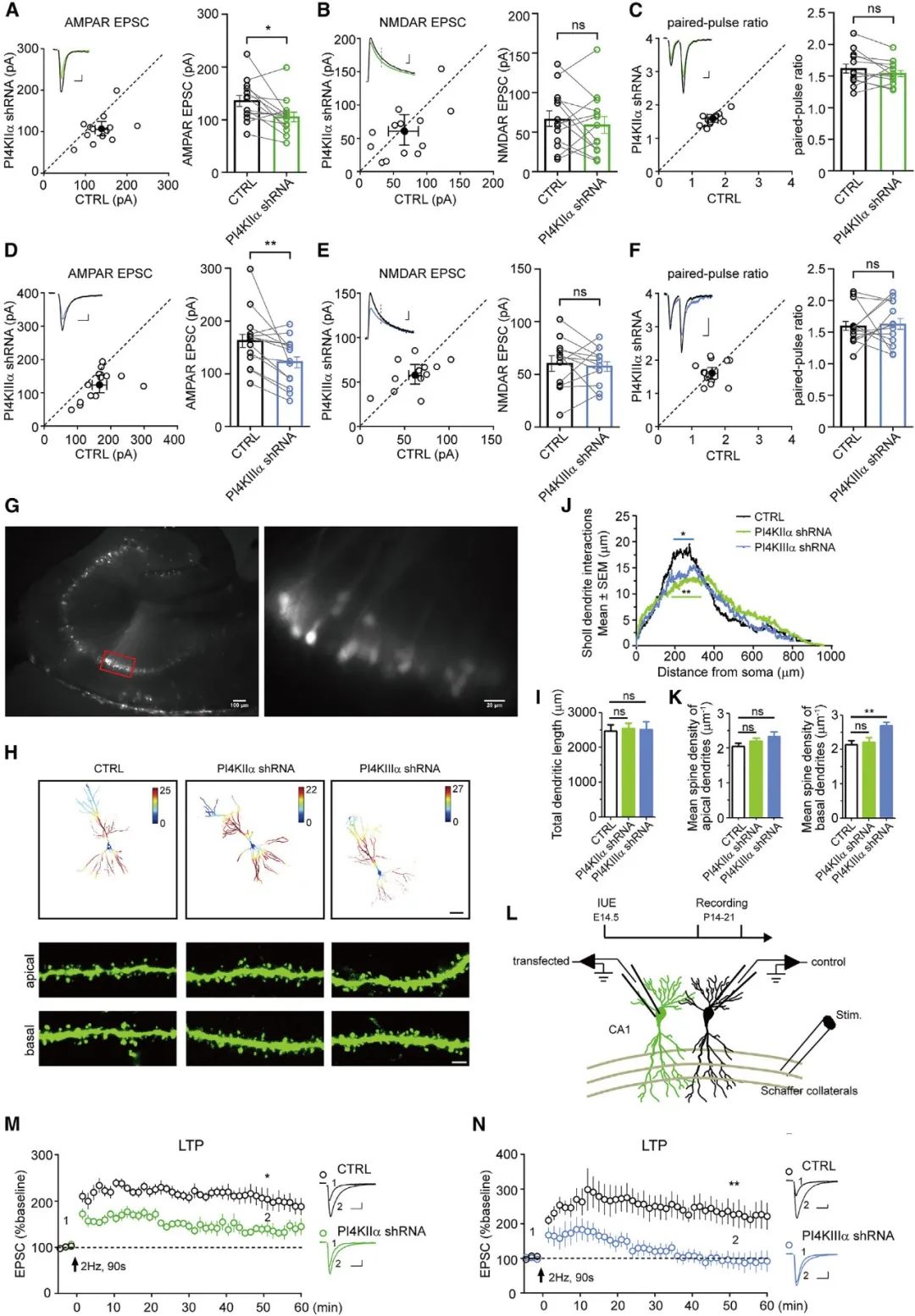
图5 沉默PI4KIIa和PI4KIIIa的表达可抑制LTP
最后,作者借助情境恐惧条件反射和Morris水迷宫行为学范式发现,在海马CA1区沉默PI4KIIIa表达可导致小鼠记忆缺陷。提示了,海马CA1神经元中PI4KIIIa依赖的PI4P合成对于学习记忆是必需的(图6)。
上述结果提示了沉默PI4KIIIa可损害了突触的长时程增强作用以及动物的记忆能力。
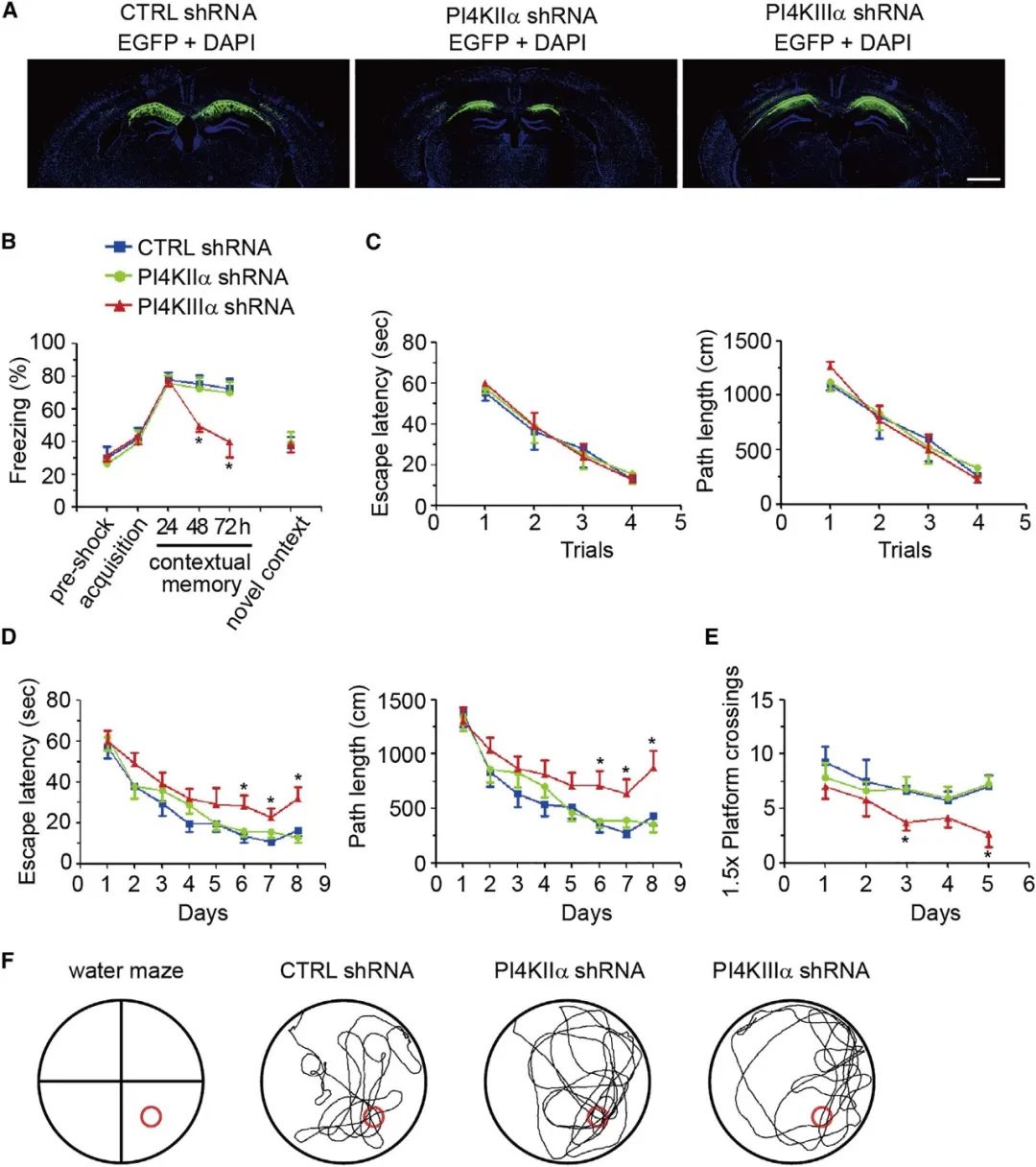
图6 沉默海马CA1兴奋性神经元中PI4KIIIa的表达导致小鼠记忆缺陷
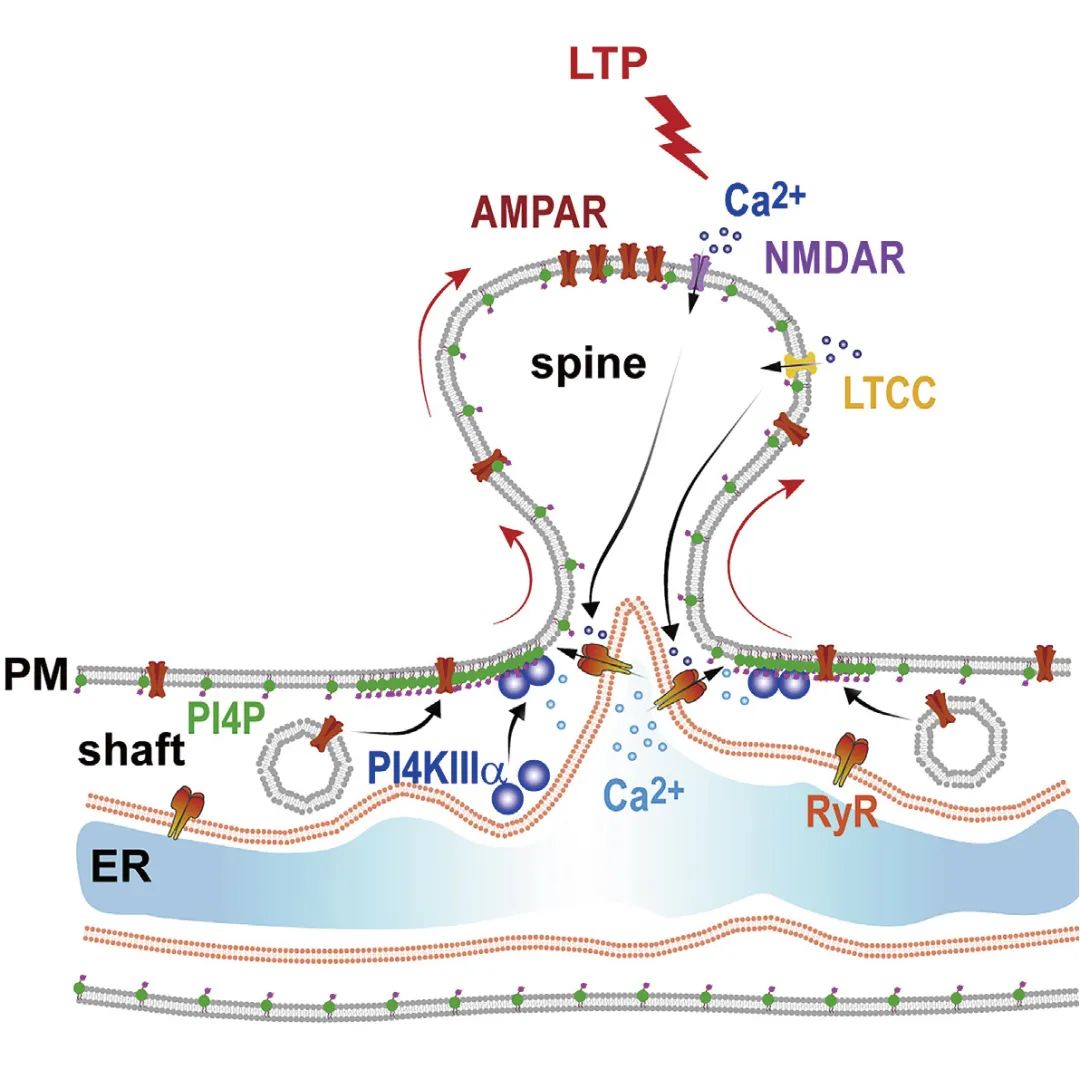
PI4P是生物膜中的低丰度磷脂分子,但具有调控脂转移和囊泡运输等重要生物学功能。刘佳佳研究员团队借助脂质组学、快速活细胞成像、细胞生物学、RNAi(RNA干扰)技术、全细胞膜片钳记录、行为学范式等多个技术手段发现小鼠海马神经元细胞质膜富含PI4P,在产生LTP的海马神经元中,细胞内钙离子浓度上升促进了PI4KIIIa定位在树突质膜内表面,使质膜PI4P水平迅速升高。进一步研究发现,通过抑制PI4KIIIa活性急性降低质膜PI4P水平可完全抑制LTP诱导期的AMPAR质膜表达,而在小鼠大脑海马CA1区沉默PI4KIIIa表达导致突触的长时程增强作用以及动物的记忆能力受损。该项研究不仅发现膜脂PI4P的代谢受到神经活性调控,而且揭示了PI4P在AMPAR运输中的调控作用以及在LTP和学习记忆中的生理意义。
刘佳佳实验室博士生郭真真和南京大学石云实验室博士后蒋朝华为该论文的共同第一作者。该研究得到了国家自然科学基金委、科技部和中国科学院战略性先导科技专项B的资助。
和元生物有幸提供实验中使用的AAV载体,用实际行动助力中国脑科学的发展。



