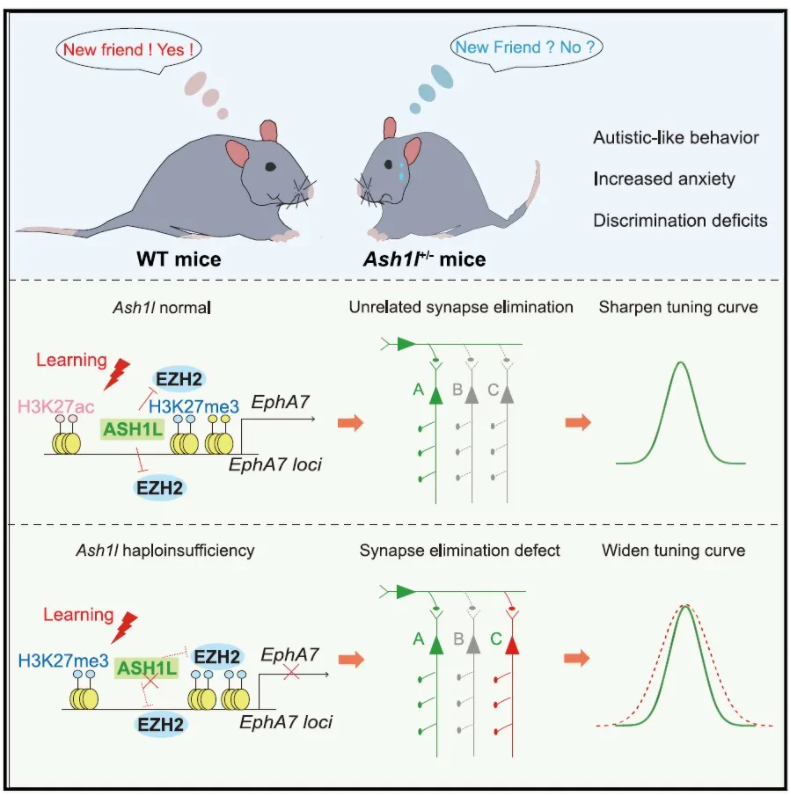
中枢神经系统的发育和学习诱导的神经环路重构都受到基因表达的时空调控。组蛋白重构在生理和病理条件下调控这类基因的表达起着重要作用,因此,组蛋白重构可能成为许多重大脑疾病的潜在药物靶点。
组蛋白甲基转移酶被确认为许多神经发育障碍的风险基因。Ash1l是一种组蛋白甲基转移酶,属于三胸家族(trxG)蛋白的成员之一,其能拮抗多梳蛋白家族对基因转录的抑制功能。目前越来越多的研究证明,Ash1l基因功能缺失是自闭症谱系障碍(autism spectrum disorder, ASD)发生的首要危险因素,然而,关于Ash1l单倍体不足导致与 ASD相关致病机制尚不清楚。
2022年1月26日,上海科技大学生命科学与技术学院管吉松教授团队利用Ash1l缺失小鼠模型、行为学范式、光遗传学等多个技术手段发现Ash1l单倍体不足导致小鼠ASD样行为,破坏活动依赖的突触修剪过程,并对该现象进行了系统性探索,相关研究成果刊发在神经科学期刊Neuron上,标题为ASH1L haploinsufficiency results in autistic-like phenotypes in mice and links Eph receptor gene to autism spectrum disorder
结果
前脑神经元Ash1l单倍体不足导致小鼠的ASD样表型
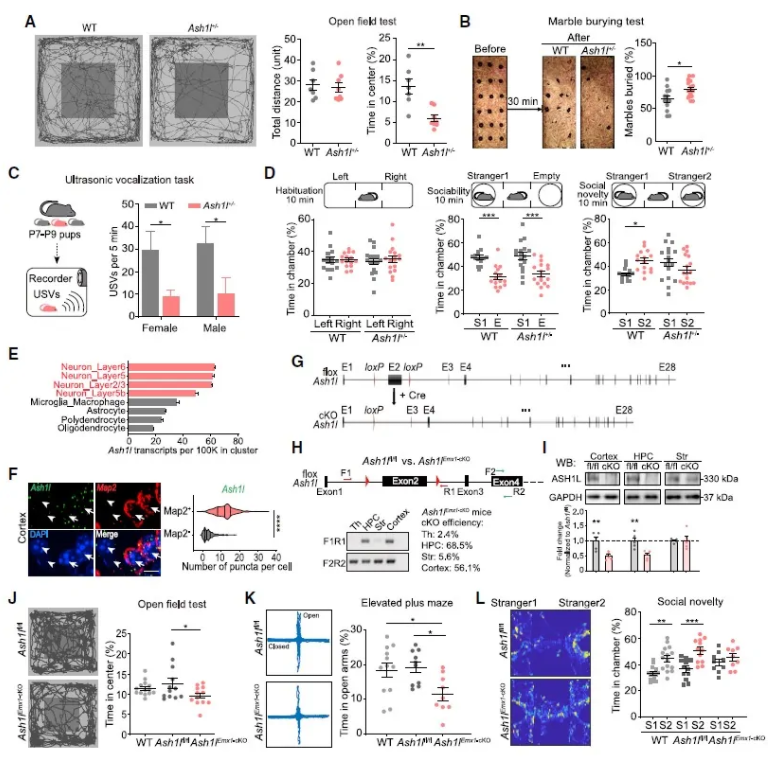
首先,作者比较了Ash1l+/-小鼠[1,2]和WT小鼠的行为学表型,在Ash1l+/-小鼠中检测到了单倍体不足,同时,旷场实验(the open-field test)和埋珠实验(the marble-burying test)行为学范式结果显示,Ash1l+/-小鼠表现类似焦虑、重复刻板的行为(图1A,B)。此外,对出生后7-9天(P7-P9)小鼠进行幼鼠超声发声测试,发现Ash1l+/-小鼠在生命早期存在现沟通障碍的现象(图1C)。三箱社交实验(three-chamber assay)结果显示Ash1l+/-小鼠也表现出社交障碍(图1D)。这些结果提示了Ash1l突变小鼠表现出典型的自闭症样行为,包括沟通困难,重复刻板行为、社交障碍等。
单细胞基因表达分析发现,在中枢神经系统中, Ash1l优先在神经元中表达(图1E)。因此,研究人员借助原位杂交链式反应(HCR-FISH)做进一步验证,发现在皮层神经元中,Ash1l阳性信号与Map2阳性信号((Map2+,神经元标志物)高度重叠(图1F);在Map2-细胞中,Ash1l信号点数量明显减少。这提示了神经元中Ash1l功能缺失可能导致自闭症谱系障碍 (Autism Spectrum Disorder, ASD)样行为。
基于上述结果,作者构建Ash1lEmx1-cKO小鼠(特异性在前脑,特别是新皮层和海马区敲除Ash1l,图1G-I),并通过行为学范式发现,前脑Ash1l单倍体不足导致小鼠ASD样行为(图1J-L)。
图1 Ash1l单倍体不足导致小鼠的ASD样表型
Ash1l突变小鼠存在辨别能力缺陷
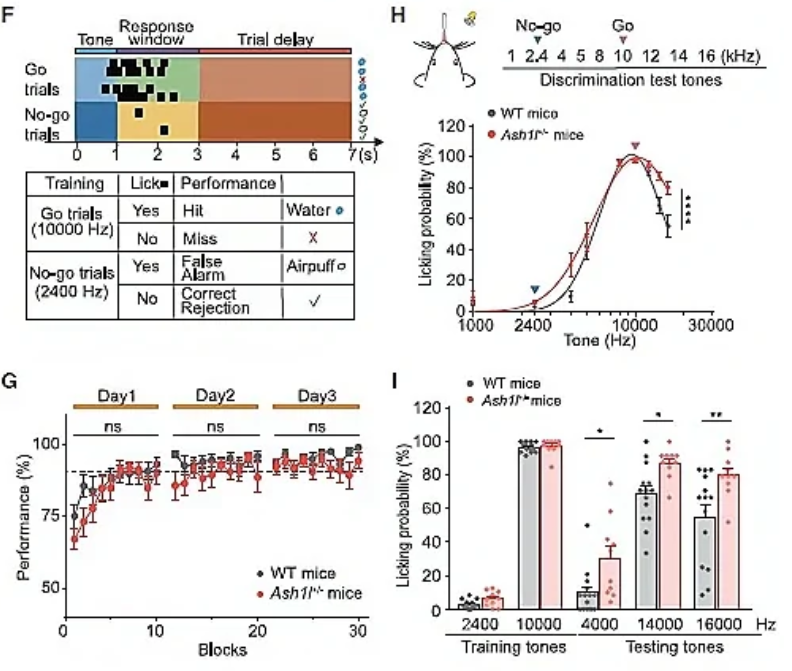
接下来,研究人员对Ash1l缺失小鼠的学习行为进行评估,借助条件性恐惧反射范式发现,Ash1l+/-小鼠和WT小鼠均学会了情境性恐惧条件反射,并表现出类似的恐惧记忆重提水平,然而Ash1l+/-小鼠在恐惧学习的第三天表现出木僵行为(freezing, 老鼠的一种防御反应)下降;此外,在Ash1l+/-小鼠中未观察到两种不同环境下的木僵行为差异(WT小鼠在context A中表现出较高的木僵行为,在context B(新环境)中表现出较低的木僵行为),提示了Ash1l+/-小鼠辨别能力受损(图2A,B)。
在音调依赖的辨别范式(图2C-E)和基于音调的Go-No-go行为范式(图2F-H)中,Ash1l+/-小鼠同样表现出明显的听觉辨别障碍(听觉脑干反应(ABR)测试Ash1l+/-小鼠无明显的听力损失)。
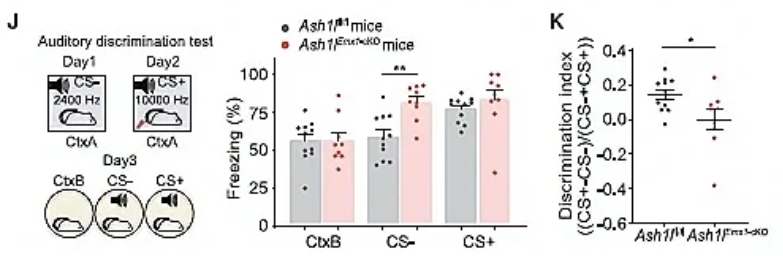
为了验证前脑神经元中Ash1l缺失是否会导致与Ash1l+/-小鼠类似的辨别能力障碍现象,作者借助Ash1lEmx1-cKO小鼠进行相关行为学范式,发现Ash1lEmx1-cKO小鼠与恐惧记忆相关的听觉辨别能力明显受损(图2J,K)。这种Ash1l突变小鼠的辨别能力异常符合部分ASD患者的临床表现。前脑Ash1l缺失可导致突触和神经环路功能障碍,以及辨别行为障碍。
图2 Ash1l+/-小鼠表现出正常的学习能力,但辨别能力受损
Ash1l以细胞自主效应的方式介导活动依赖的突触修剪过程
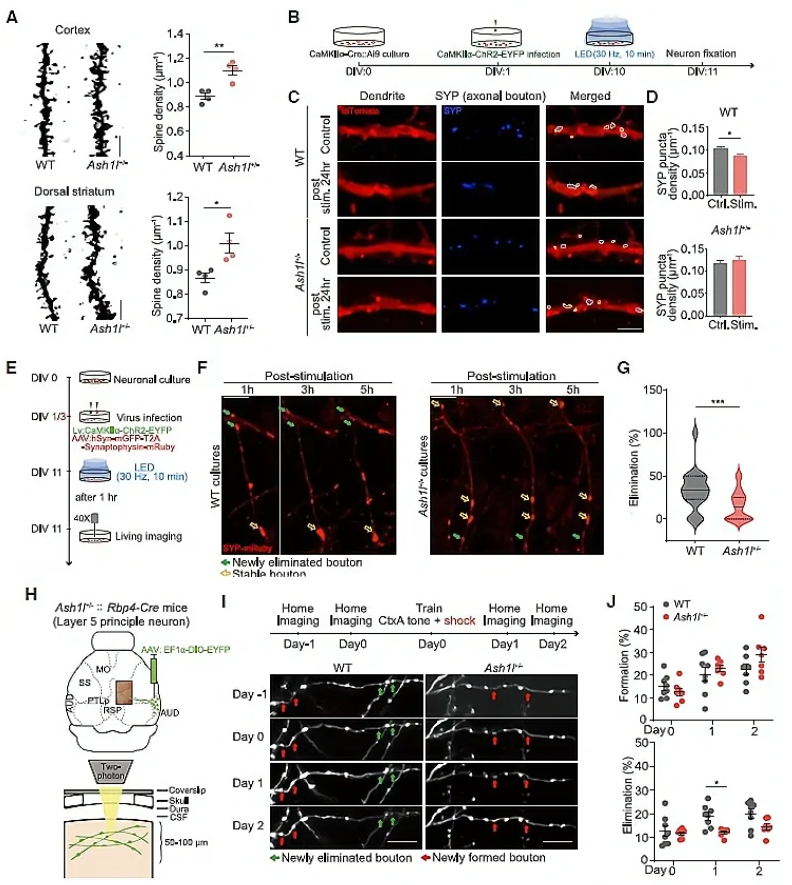
随后,作者借助高尔基染色发现,1月龄的Ash1l+/-小鼠皮层神经元和背侧纹状体中棘神经元(MSNs)的树突棘密度显著增加(图3A),为研究这种现象是否源于突触消除过程(synapse elimination)的减少,研究人员研究了Ash1+/-小鼠原代皮层神经元的突触密度变化。用表达ChR2光敏蛋白的慢病毒载体(Lenti-CaMKIIa-ChR2)感染培养的神经元,并用LED刺激(图3B),发现WT神经元在LED刺激后24h树突突触素(SYP)密度显著降低,而Ash1l+/-组则未观察到这种现象(图3C,D),提示了Ash1l可能介导了活动依赖的突触修剪过程。活体成像结果显示,Ash1l+/-组培养的神经元中,活性依赖的突触消除过程被削弱。
此外,作者还研究了在小鼠新皮层中活性依赖的突触消除,追踪了听觉皮层(auditory cortex, AUD)到后顶叶皮层(posterior parietal cortex, PTLp)长距离投射轴突的变化。研究人员将AAV-EF1a-DIO-EYFP注射在Rbp4-Cre小鼠AUD,特异性标记听觉皮层L5神经元(图3H)。小鼠接受音调依赖的恐惧条件训练,成像观察轴突变化发现,声调依赖的恐惧条件学习24h后,Ash1l+/-::Rbp4-Cre小鼠“bouton”(突触小结)的清除明显低于WT小鼠,而“bouton”形成则保持不变(图3H-J)。
为探究这种“突触修剪”过程的缺失是否由Ash1l缺失神经元的细胞自主效应导致的,研究人员将AAV-hSyn-Cre和AAV-EF1a-DIO-EYFP注射于Ash1lfl/fl小鼠右侧AUD区(图3K),对小鼠进行音调依赖的恐惧条件行为学范式训练,在学习之后借助体内成像技术追踪突触的变化,发现Ash1l缺失的神经元中学习依赖的突触的增加数量没有发生改变,而学习诱发的突触的减少数量显著低于正常小鼠(图3K-M)。这些数据提示了Ash1l以细胞自主效应的方式介导皮层神经元的活动依赖的突触修剪过程。
图3 Ash1l以细胞自主效应的方式介导活动依赖的突触修剪过程
Ash1l调控幼鼠和成年小鼠中EphA7的表达
接下来,作者研究了其分子机制,通过对转录组分析及基因本体论(GO)分析发现,Eph受体EphA7在听觉皮层和背侧纹状体中的表达均显著下调(图4A-C),基于此,研究人员进一步将EphA7作为参与Ash1l介导的突触修剪过程的重要基因之一进行了研究。
Eph受体及其配体 Ephrin 统称为Eph家族蛋白,是蛋白酪氨酸激酶家族中的最大成员。Eph/ephrin蛋白作为细胞表面分子,参与调节突触的形成及神经元的可塑性,EphA7是编码Eph受体中14个受体蛋白基因之一,在皮质发育和突触功能中发挥重要作用。
随后,研究人员借助音调依赖的恐惧条件行为学范式,在体内/体外均发现Ash1l+/-小鼠听觉皮层活动依赖的神经元中EphA7表达远低于WT小鼠的(图4D-I),提示了在Ash1l+/-小鼠中EphA7活动诱导表达受损。
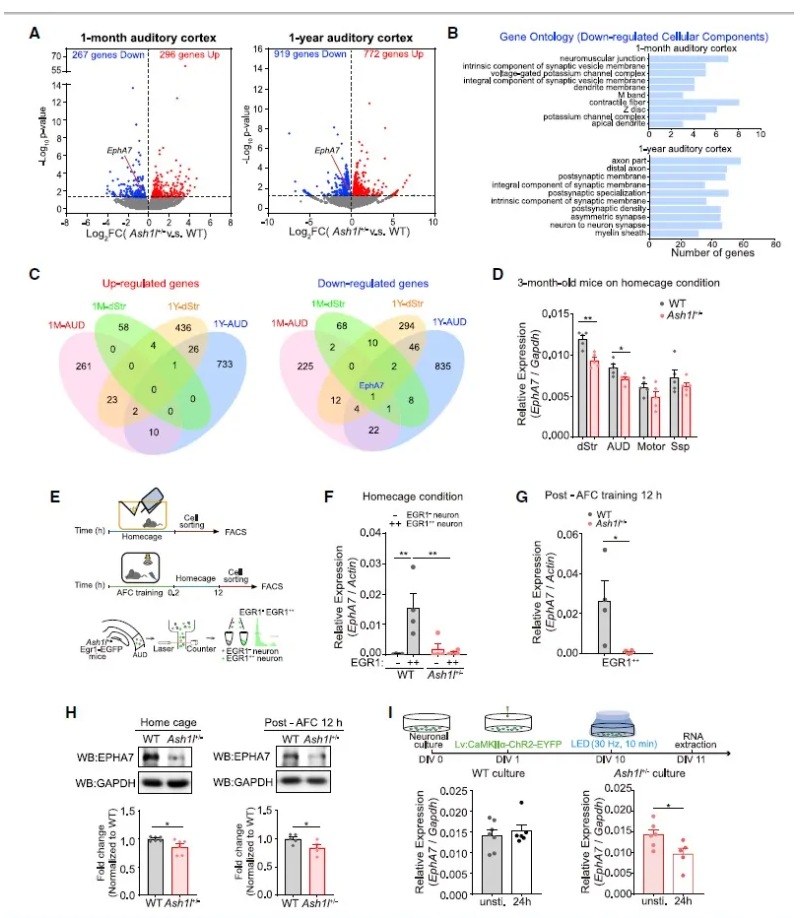
图4 Ash1l调控幼鼠和成年小鼠中EphA7的表达
Ash1l通过拮抗EZH2-H3K27me3来调控EphA7的表达
进一步,作者探究Ash1l是如何参与调控EphA7活动诱导表达,通过染色质免疫沉淀技术(ChIP)发现,Ash1l的直接靶点H3K36me2在WT和Ash1l+/-小鼠AUD EphA7基因位点附近没有明显差异,然而H3K27me3信号(组蛋白甲基转移酶复合体PRC2催化的抑制性标记物)在Ash1l+/-小鼠EphA7基因位点显著增加(图5A,B)。同时,Ash1l+/-小鼠中H3K27ac(一个活跃的增强标记。H3K27ac与H3K27me3共享一个位置,存在拮抗作用。)表达持续下降,用NaB(组蛋白去乙酰化酶抑制剂)预处理培养的神经元发现,刺激Ash1l+/-神经元后H3K27ac升高,EphA7表达恢复(图5C,D)。同样地,行为学结果显示,Ash1l+/-小鼠中H3K27me3、EZH2(PRC2复合体的一个重要的催化酶)表达水平显著增加(图5E-G)。通过UNC1999(EZH1/2抑制剂)对培养的神经元进行预处理,发现Ash1l+/-激活的神经元中EphA7表达受到抑制的现象得到改善(图5I)。上述结果提示了,Ash1l在活性依赖的PRC2介导的转录抑制累积中发挥拮抗作用,从而允许EphA7在激活的神经元中表达。

图5 Ash1l通过拮抗EZH2-H3K27me3沉默来调控EphA7的表达
EphA7高表达可改善Ash1l+/-小鼠突触修剪过程缺陷和行为异常
基于上述结果,研究人员研究了提高EphA7受体活性是否能改善Ash1l突变小鼠的突触修剪过程缺陷和行为异常。Ephrin-A5是EphA7的高亲和力配体,参与下游信号[3,4]。作者在培养的神经元和小鼠听觉皮层中应用Ephrin-A5多聚体的Fc融合蛋白(Ephrin-A5 Fc)直接刺激EphA7活性[5,6],发现改善了Ash1l突变小鼠的突触修剪过程和行为异常(图6)。
图6 Ephrin-A5激活EphA7高表达Ash1l+/-小鼠突触修剪过程缺陷和行为异常
结论
本文借助转基因模型鼠、多种行为学范式、光遗传学技术、活体成像转录组分析及基因本体论分析等多种技术手段,研究发现前脑神经元Ash1l单倍体不足可导致小鼠产生沟通困难、重复刻板行为、社交障碍等类似自闭症样表型,并机制上破坏活动依赖的突触修剪过程,这是以细胞自主效应方式介导的。进一步研究发现,Ash1l可通过介导EZH2-H3K27me3去抑制作用提高EphA7的活性依赖表达,进而影响突触消除过程,并改善Ash1l+/-小鼠辨别能力障碍及行为障碍。该研究有助于深入了解ASD相关发病机制,为其干预治疗提供了新的思路。
作者介绍及基金信息
正在上科大管吉松教授课题组进行合作研究的2017级博士研究生闫宇泽(清华大学)和上科大生命学院的田苗苗博士为本文的共同第一作者。上科大生命学院研究生 2016级陈琪楠、2017级郭修贤、2018级李萌、2019级周罡也对本研究做出了重要贡献。管吉松教授、青岛大学附属医院刘世国教授和清华大学熊巍教授为本文的共同通讯作者,研究得到中国科学院遗传发育所吴青峰研究组、上海理工大学谢红研究组的大力支持。研究也得到了上科大戚炜教授、范高峰教授和上海交通大学医学院徐楠杰教授大量的有益帮助。
该研究受到科技创新2030-重大项目资助,和自然科学基金项目以及上海市科委项目支持。
和元生物有幸提供实验中使用的AAV、慢病毒载体,用实际行动助力中国脑科学的发展。
参考文献
[1] Liu, S.G., Tian, M.M., He, F., Li, J.N., Xie, H., Liu, W.M., Zhang, Y.T., Zhang, R., Yi, M.J.,Che, F.Y., et al. (2020). Mutations in ASH1L confer susceptibility to Tourette syndrome. Mol Psychiatr 25, 476-490.
[2] Zhu, T., Liang, C., Li, D., Tian, M., Liu, S., Gao, G., and Guan, J.S. (2016). Histone methyltransferase Ash1L mediates activity-dependent repression of neurexin-1alpha. Sci Rep 6, 26597.
[3] Beuter, S., Ardi, Z., Horovitz, O., Wuchter, J., Keller, S., Saha, R., Tripathi, K., Anunu, R., Kehat, O., Kriebel, M., et al. (2016). Sci Rep 6,1451 29710.
[4] Nguyen, T.M., Arthur, A., Zannettino, A.C.W., and Gronthos, S. (2017). EphA5 and EphA7 forward signaling enhances human hematopoietic stem and progenitor cell maintenance,migration, and adhesion via Rac 1 activation. Exp Hematol 48, 72-78.
[5] Clifford, M.A., Athar, W., Leonard, C.E., Russo, A., Sampognaro, P.J., Van der Goes, M.S., Burton, D.A., Zhao, X.M., Lalchandani, R.R., Sahin, M., et al. (2014b). EphA7 signaling guides cortical dendritic development and spine maturation. P Natl Acad Sci USA 111, 4994-4999.
[6] Depaepe, V., Suarez-Gonzalez, N., Dufour, A., Passante, L., Gorski, J.A., Jones, K.R., Ledent, C., and Vanderhaeghen, P. (2005). Ephrin signalling controls brain size by regulating apoptosis of neural progenitors. Nature 435, 1244-1250.